人种
人种(race),具有共同遗传体质特征的人类群体。人种也称种族,在生物学上,人类各种族都同属于一个物种,即智人。不同的种族相当于在一个物种下的若干变种,他们都起源于一共同祖先。不同的人种虽然在肤色、眼色、发色、发型、头型、身高等特征上有所区别,但这些特征差异是由于人类在一定地域内长期适应当地自然环境,又经长期隔离所形成的。
分类 根据体质特征的差异,过去全世界人种大致上分为3大人种,即蒙古人种(黄色人种)、高加索人种(白色人种)、尼格罗人种(黑色人种)。有人还主张再分出澳大利亚人种(棕色人种)来。在这些主要人种之间还有若干过渡人种,如介于尼格罗人种和高加索人种之间的有埃塞俄比亚人种和南印度人种,介于高加索人种和蒙古人种之间的有南西伯利亚人种和乌拉尔人种,介于蒙古人种和尼格罗人种之间的有波利尼西亚人种和千岛人种。随着人类历史的发展,人类的生产活动能力的提高,控制自然能力的增强,社会因素逐渐加强而自然选择因素逐渐削弱,人类交往日益频繁,各人类集团间不断发生混杂,混杂现象越来越广泛,几乎扩展到了世界上每个角落,因此,世界上几乎没有一个绝对的纯种(图1)。
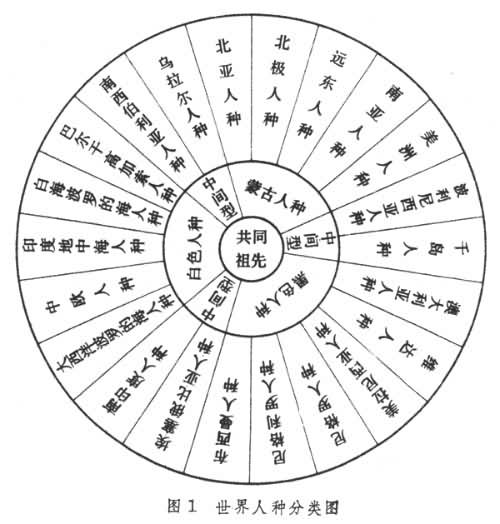
现代各人种的分布 除去各种水域和一些高山外,人类是在地球上分布最广的一种生物。
现代人的分布经历了一个相当长时期的发展,至少可以追溯到更新世晚期的后半部,即旧石器时代晚期的晚期智人阶段。晚期智人具有高而圆隆的头颅和相对较小的面部,低矮的眉弓和突出的下颏等特征,它是现代智人的一种早期类型。在地球上广泛分布的最早的晚期智人各群体已经积累了文化上和体质遗传上的差异,并在此后3.5万年以上的时间里进一步得到扩展和改变,形成了现生各个人种的特征和分布状况。
与动物物种产生和分化的过程一样,形成现代各人种的自然过程仍然受基因突变、变异的随机扩散、迁移和自然选择四个基本因素控制。在这些因素作用下,现生各人种在某些形态特征和某些生物化学特征上显示了程度不同的差异,上述生物化学特征包括ABO、Rh、MN等多种血型系统和其他血液因子系统,某些遗传性疾病的存在或出现频率,以及对某些疾病的遗传性抗御等等。
各人种的体质形态一般来说与他们的生活环境是相适应的。例如,居住在炎热的非洲沙漠的尼格罗人的肢体细长;而在严寒的北极生活的因纽特人(以前一般称为“爱斯基摩人”)则肢体粗短。这与身体热量的保持和散发有关。赤道附近尼格罗人的皮肤含有较多的色素,可以避免过多的紫外线照射,他们卷曲的头发也可起隔热的作用。蒙古人的倾斜的凤眼和内眦褶,还有宽的和富含脂肪的面颊、较平的鼻部和额部,可能与亚洲中部寒冷的多风沙气候有关。
在人种形成的过程中,自然因素作用的大小、性质和诱发条件,随着人类物质生产和社会文化的发展而变得越来越不同于其他动物物种形成的过程。人种的形成和分布比在其他动物同一物种内不同亚种的情况复杂得多。因此,不能简单地用自然条件来解释某些人种的某些特征。虽然非洲尼格罗人皮肤黑色素的骤增与太阳照射有关,但是实际上现在的非洲尼格罗人大部居住在蔽荫的森林里。世界上个子最高的人是居住在苏丹尼罗河上游的尼罗特人,成年男子平均身高达180厘米。有趣的是,世界上最矮的俾格米人也居住在这个区域。斯堪的纳维亚半岛低矮的拉普人也与高大的北欧人种毗邻居住,非洲尼格罗人面部突颌的特征适宜于炎热的气候,但北极人种中的阿留申类型的突颌比尼格罗人还要明显。这样一些表面上与自然条件相矛盾的人种特征的形成过程更复杂一些。
各人种的起源 蒙古人种、高加索人种、尼格罗人种三大人种主干的体质特征是在约 3.5万年前开始出现的。在亚、欧、非三大洲发现的晚期智人的化石证明他们在本地区延续发展。美洲、澳洲和大洋洲较晚才有人类居住。由于澳洲土著的体质特征较为特殊,他们常被列为与以上三种人不同的另一种人。
蒙古人种起源于中亚和东亚。中国北京周口店山顶洞和广西柳江县通天岩发现的晚期智人头骨化石表现了以黄种人特征为主导的特征。在马来西亚沙捞越的尼亚洞和在印度尼西亚爪哇瓦贾克发现的人类化石很像现代澳大利亚土著。早期蒙古人种逐渐向北方扩展,成为西伯利亚的通古斯人和楚克奇人以及因纽特人(爱斯基摩人)。跨过西伯利亚和美洲阿拉斯加之间曾经存在的“陆桥”,蒙古人种来到美洲,定居下来,成为美洲大陆最早的居民──印第安人。
蒙古人种从印度尼西亚,或者还有其他的地方,航海到澳大利亚,比到美洲去的历史或许稍早一些。在澳大利亚的芒戈湖、科阿沼泽发现的人类化石代表了两种不同的类型,因此有人认为曾经有过两次从亚洲向澳洲的迁移。大洋洲的美拉尼西亚、波利尼西亚和密克罗尼西亚可能只是在几千年前才有人居住。
非洲尼格罗人与在本地区发现的晚期智人在体质特征上的继承关系很明显,大部分人认为他们起源于非洲本土。但是,地区内变种分化的较晚。南非和东非的晚期智人化石都在不同程度上包含尼格罗人 (Negroes)、科伊桑人(Khoisans,即以前所称的“布须曼人和霍屯督人”)和当地现代人特征的混合。只是在几千年之前,科伊桑人才在非洲南部定居下来,在北部,与狩猎经济相适应,形成尼格罗类形。
与欧洲晚期智人有关系的早期智人,即尼安德特人(Neandertals) 阶段的化石比较复杂。最早的白种人的起源地目前只能圈定在南欧、北非和西亚这样一块较大的区域之中。欧洲本土发现的晚期智人化石以克罗马农人(Cro-Magnon)为代表,他们在3万年前已经显示了高加索人种的性质。由于欧洲与附近亚、非洲部分的人群混杂很频繁,高加索人种再分化为现代的次级人种则是大约 1万年以内的事。这样的次级人种包括波罗的海周围浅肤色的北欧人种、浅黑色皮肤的地中海人种和北非埃塞俄比亚人种以及深褐色皮肤的南印度人种。1000多年以前宗教对婚姻的限制也是后期人种特征相对稳定的重要因素。
意大利航海家C.哥伦布1492年到达美洲大陆以后,随着欧洲资本主义国家寻求新的殖民地,世界人种的分布发生了急剧的变化:数以百万计的欧洲人涌向美洲、南非、澳大利亚和新西兰,使高加索人种的数目迅速增长。例如,近4个世纪以来世界人口增长约6倍,而英国人的后裔至少增长50倍。与此同时,几百万非洲黑人作为奴隶被绑架到美洲。由于混杂又出现了新的人种,美洲的有色人种已达6000万人之多。有的人种(塔斯马尼亚人)已灭绝,有些人种如澳洲土著及美洲印第安人人口大大减少。这样的一系列过程形成了现代人种分布的基本格局(图2)。
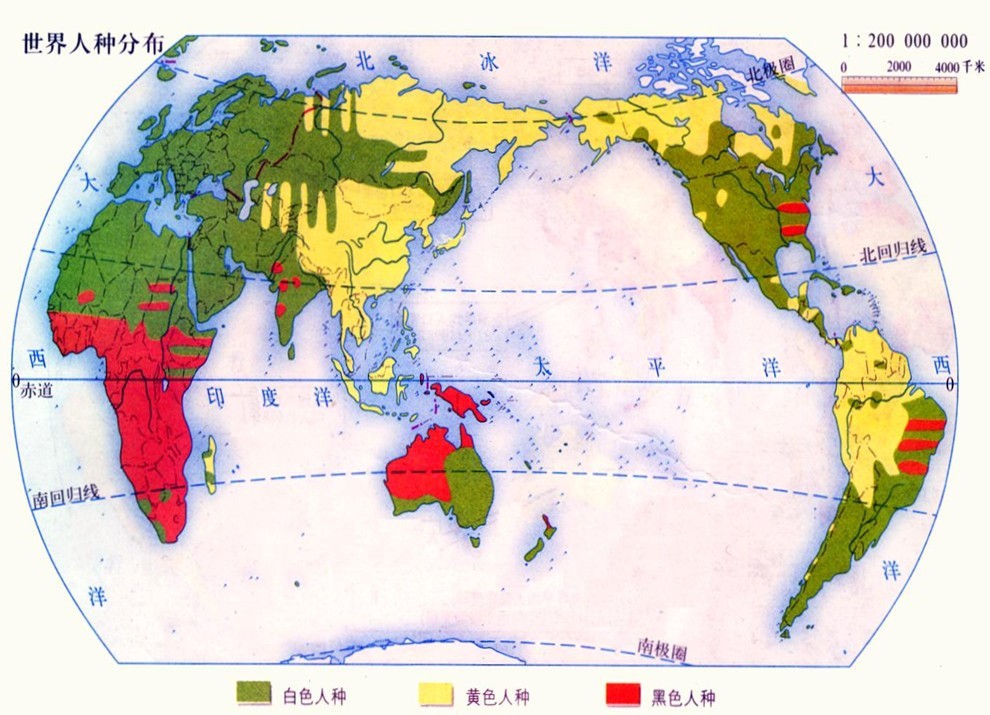
人种划分的补充说明 除外表形态特征外,在生物化学及人类遗传学研究的基础上,发现过去以纯形态学为标准的三分或四分的人种划分方案已经不能全面反映真实情况了。至于完全以肤色为标准的“白种人”、“黄种人”、“黑种人”、“棕种人”,甚至还有“红种人”的名称也并不十分恰当,应该摒弃。
基于科学的发展,在外表形态特征基础上,加上血型、遗传病等的差异,再考虑地理阻障等因素,把世界人种划分为9个地理人种(geographical race)。
印度地理人种的外表特征虽与高加索人种有许多相同之处,但肤色较深,身材细长,有些属骨瘦如柴类型。此外,骨密度低;B型血型出现率高,Rh阴性血型出现率低。显示其血型特征上与蒙古地理人种相似。
高加索地理人种的Rh阴性血型出现率高,并有A2型血型的人,与印度地理人种不同。其肤色浅、体毛浓密。鼻大而窄和男子秃顶者多也是其显著特征。
印第安地理人种或称美洲印第安地理人种,过去曾依外表特征而归于蒙古人种,如直黑发、铲形门齿出现率高、体毛稀疏、男子秃顶者少等。但其肤色较蒙古人种更浓重,且具偏红色调。其血型特征也与蒙古人种不同,如无B型及Rh阴性血型的人,N型血型的人也很少,而Diego阳性血型出现率高。
蒙古地理人种在血型特征上是 B型血型出现率高,Duffy血型及Diego阳性血型出现率不稳定或偏高。
尼格罗地理人种从外形特征上有时可以划分为具典型特征的刚果人种和肤色微黄,身材矮细、具螺旋状丛生发的开普人种(包括科伊桑人)。二者在血型上无明显差异。一般R0型血型出现率高,U-阴性血型略少,Fy型和V型血型很罕见。血红蛋白异常。有6-磷酸葡萄糖脱氢酶缺乏症。
澳大利亚地理人种肤色浓黑,发波状至卷曲状,身材细长、体毛中等至浓密、男子秃顶率高、并伴有小腿无毛现象。牙齿大、眉嵴高、下颌突出。B型血型出现率很低,N型血型出现率极高,无S型血型。无异常血红蛋白,无6-磷酸葡萄糖脱氢酶缺乏症,与尼格罗人种相反。
大洋洲诸岛上的人种曾与澳洲人种共称澳洲大洋洲人种。美拉尼西亚地理人种外貌上近似澳大利亚人种,但铲形门齿出现率高,B型和S型血型出现率高。有6-磷酸葡萄糖脱氢酶缺乏症和地中海贫血基因。可能澳大利亚人种与之关系更密切些。
密克罗尼西亚地理人种曾被认为可能是东南亚蒙古人种、美拉尼西亚人种和波利尼西亚人种的混血后裔。但其B型血型出现率稍高、N型较M型血型多、Duffy-阳性和Diego-阴性血型出现率很高。表明他们是独立的地理人种。
波利尼西亚地理人种肤色较浅、体毛少。B型血型出现率低、N型血型出现率较高,无Rh-阴性和A2型血型,B型、M型和Duffy-阳性血型出现率也不高。虽然面部特征近似蒙古人种;但肤色浅,波状发和浓密络腮胡又与高加索人种近似。其血型与蒙古人种及高加索人种均有较大差异。
在地理人种范围内,基于生物学的或其他如地理阻障、社会、民俗等因素,甚至历史和政治因素等而形成一些稍小的居群集团,可以称之为地域人种(locol race)。他们往往只在本群内通婚,极少与其他集团交往。一个地域人种内的各成员间,生物学特征极相似。往往许多在遗传特征上相似的地域人种可以在一个大陆或某一较大区域构成地理人种。处于边缘地区、地理阻隔严重、人口很少的地域人种常常构成在遗传性质上稳定的特殊的居群。
在人口稠密或地理阻限不严重的经济发达地区,则诸多地域人种间生物学上的差异较少,而政治、历史等方面的差异起作用较大。例如高加索人种中就有阿尔卑斯、地中海、东北欧、西北欧等地域人种。日本的阿伊努人、非洲的俾格米人、夏威夷群岛的夏威夷人都属于特定的地域人种。
在人数较多,人口密度较大而又少流动的地方常有小的居群集团;他们常表现为具单独的遗传因素,目前名之为少数人种 (microrace)。斯堪的纳维亚的拉普人可作为少数人种的例子。
人类的种族(Races of Mankind)这一概念使用已久,目前常把人种写成(human population),也就是赋予人种以居群意义。
人种的起源、形成、分布和混杂等问题至今仍有许多地方无法弄清。有待今后深入研究。